
Bacillus thuringiensis (Bt) là một loại vi khuẩn có độc tố giống với một số hợp chất hóa học được sử dụng thương mại để kiểm soát côn trùng trong sản xuất nông nghiệp và sức khỏe cộng đồng. Điều quan trọng là Bt an toàn cho con người và là thuốc trừ sâu sinh học tương thích với môi trường được sử dụng rộng rãi nhất trên toàn thế giới. Hơn nữa, gen Bt diệt côn trùng đã được tích hợp vào một số loại cây trồng chính, làm cho chúng có khả năng kháng côn trùng và do đó cung cấp một mô hình cho kỹ thuật di truyền trong nông nghiệp.
Lịch sử của vi khuẩn Bacillus thuringiensis
Vi khuẩn Bt được tìm thấy vào năm 1901, một nhà khoa học Nhật Bản tên là Shigetane Ishiwata đã phân lập được một loại vi khuẩn từ ấu trùng tằm chết trong khi ông đang điều tra nguyên nhân của căn bệnh được gọi là “bệnh sotto” (bệnh đột ngột suy sụp). Căn bệnh này là nguyên nhân dẫn đến việc mất một số lượng lớn tằm ở Nhật Bản và các vùng lân cận. Ishiwata đặt tên vi khuẩn là Bacillus sotto. Vài năm sau đó, vào năm 1911, một nhà khoa học người Đức Ernst Berliner đã phân lập được một chủng có liên quan từ những ấu trùng bột mì đã chết ở Địa Trung Hải mà ông tìm thấy trong một nhà máy bột ở bang Thuringia của Đức. Ông đặt tên thích hợp cho sinh vật là Bacillus thuringiensis. Berliner đã nghiên cứu vi khuẩn và tìm thấy các thể bao gồm “Restkorper” cùng với nội bào tử. Vào nằm 1927, Mattes một lần nữa quan sát thấy các vật thể bao gồm tương tự ở Bt nhưng mãi đến sau đó (25 năm), hoạt động diệt côn trùng mới được cho là do những vật thể có độ mềm dẻo cao này ngày nay được gọi là “tinh thể parasporal,” cụm từ được đặt ra bởi Christopher Hannay vào năm 1953. Khi Thomas Angus nhận ra tầm quan trọng của các tinh thể parasporal, ông đã nhanh chóng chứng minh trong cùng năm hoạt động diệt côn trùng của các vật thể này. Và cùng với Philip Fitz-James, Hannay vào năm 1955 đã phát hiện ra rằng các tinh thể parasporal độc hại được cấu tạo từ protein.
Thuốc trừ sâu thương mại đầu tiên dựa trên Bt, Sporine, được sản xuất tại Pháp vào năm 1938 và được sử dụng chủ yếu để kiểm soát sâu bướm bột. Tại Hoa Kỳ, Bt lần đầu tiên được sản xuất thương mại vào năm 1958 và đến năm 1961, chất diệt khuẩn sinh học dựa trên Bt đã được đăng ký bởi Cơ quan Bảo vệ Môi trường Hoa Kỳ. Kể từ năm 1996, cây trồng chuyển gen kháng côn trùng, được gọi là cây trồng Bt, đã mở rộng trên toàn cầu và đang tỏ ra khá hiệu quả và hữu ích trong việc giảm thiểu việc sử dụng thuốc trừ sâu hóa học. Các ước tính mới nhất chỉ ra rằng hơn 50% bông và 40% ngô trồng ở Mỹ được biến đổi gen để tạo ra độc tố diệt côn trùng Bt. Thị trường thuốc trừ sâu toàn cầu hiện nay (thuốc diệt cỏ, thuốc trừ sâu, thuốc diệt nấm, thuốc diệt nấm mốc và thuốc khử trùng) trị giá 25,3 tỷ USD. Thuốc trừ sâu sinh học chỉ chiếm 2,5% thị trường này nhưng thị phần của chúng dự kiến sẽ tăng lên khoảng 4,2%, tương đương hơn 1 tỷ USD, vào năm 2010.
Điều thú vị là một số chủng Bt tạo ra các protein không có tác dụng diệt côn trùng mà kết tinh thành các thể vùi parasporal có hình dạng bất thường. Sự pha trộn của một chủng phân lập được xử lý bằng protease độc đối với các tế bào ung thư ở người, bao gồm tế bào ung thư bạch cầu T (MOLT-4) và tế bào ung thư cổ tử cung (HeLa).
Vòng đời của Bacillus thuringiensis
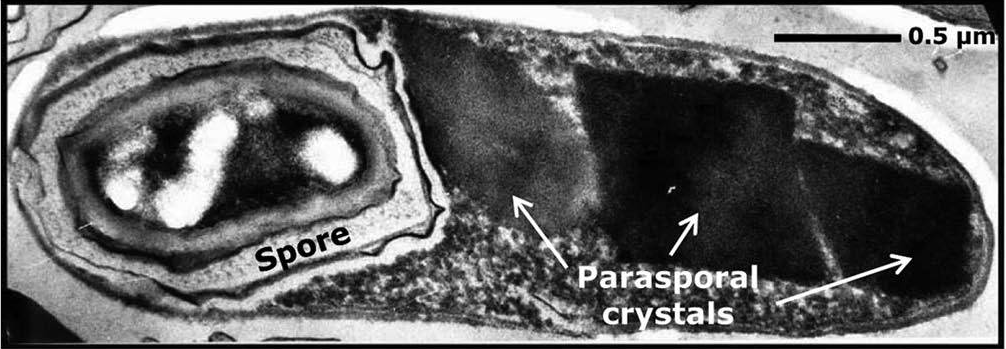
Chu kỳ sống của Bt được đặc trưng bởi hai giai đoạn bao gồm phân chia tế bào sinh dưỡng và phát triển bào tử, hay được gọi là chu kỳ bào tử. Tế bào sinh dưỡng có dạng hình que (dài 2–5 µm và rộng khoảng 1,0 µm) và phân chia thành hai tế bào con đồng nhất bằng cách hình thành vách ngăn phân chia bắt đầu từ giữa dọc theo màng sinh chất. Mặt khác, bào tử liên quan đến sự phân chia tế bào không đối xứng và được đặc trưng bởi bảy giai đoạn bao gồm (giai đoạn I) hình thành sợi trục, (giai đoạn II) hình thành vách ngăn tiền bào tử, (giai đoạn III) sự kết dính, sự xuất hiện lần đầu tiên của các tinh thể parasporal và hình thành tiền bào tử, (giai đoạn IV đến VI) sự hình thành khoang ngoài, thành tế bào nguyên thủy, vỏ não và lớp áo bào tử kèm theo sự biến đổi của nucleoid bào tử và (giai đoạn VII) trưởng thành bào tử và ly giải bào tử. Hình 1 mô tả một tế bào Bt đã hình thành hoàn chỉnh, trong đó có một số tinh thể parasporal nằm dọc theo nội bào tử. Việc sản xuất các protein tinh thể bởi Bt trong quá trình bào tử là một hiện tượng sinh học được điều chỉnh về mặt di truyền độc đáo, có lẽ, làm giảm căng thẳng về thể chất bằng cách bù đắp lượng nước mất đi trong quá trình hình thành bào tử và tạo thêm lợi thế sống sót bằng cách gây chết người đối với côn trùng vật chủ.
Phân loại Bacillus thuringiensis
Như đã chỉ ra ở trên, các tế bào sinh dưỡng của Bt có đặc điểm là hình que lớn, thẳng hoặc hơi cong với các đầu tròn. Chúng thường xảy ra theo cặp hoặc chuỗi ngắn. Bt là Grampositive, không hình mũ và di động bằng roi phúc mạc. Việc phân loại các chủng Bt đã được thực hiện bằng cách phân loại huyết thanh H, phản ứng miễn dịch học với kháng nguyên trùng roi của vi khuẩn. Trình tự axit amin của roi cụ thể có tương quan với các kiểu huyết thanh Bt H cụ thể và ít nhất 69 kiểu huyết thanh H và 82 kiểu huyết thanh học (serovars) của Bt đã được đặc trưng. Tuy nhiên, việc phân loại huyết thanh H bị hạn chế ở khả năng phân biệt các chủng từ cùng một kiểu huyết thanh H hoặc từ cùng một serovar. Do tầm quan trọng về kinh tế của nó, việc phát triển các công cụ thay thế để phân loại và phân nhóm các chủng và chủng phân lập Bt trở nên cần thiết. Do đó, một số chương trình sàng lọc đã được thiết lập để phân lập các chủng Bt mới với các đặc tính diệt côn trùng độc đáo. Kết quả là, nhiều chủng Bt có hoạt tính chống lại côn trùng lepidopteran, dipteran và coleopteran đã được phân lập. Ngoài ra, các chủng Bt hoạt động chống lại côn trùng thuộc các bộ Hymenoptera, Homoptera, Orthoptera và Mallophaga cũng như tuyến trùng, ve và động vật nguyên sinh đã được phân lập.
Việc đặt Bt như một loài riêng biệt trong chi Bacillus đã gây tranh cãi kể từ khi xuất bản Chi Bacillus năm 1973, và Sổ tay về vi khuẩn học xác định của Bergey năm 1974. Chi Bacillus là một trong những chi đa dạng nhất trong lớp Bacilli và bao gồm các vi khuẩn hình thành bào tử Gram dương hiếu khí và kỵ khí đa dạng, hình que, với hàm lượng G + C dao động từ 32-69%. Dựa trên sự không đồng nhất về phát sinh loài, tám chi trong nhóm Bacilli đã được đề xuất: Bacillus, Alicyclobacillus, Paenibacillus, Brevibacillus, Aneurinibacillus, Virgibacillus, Salibacillus và Gracilibacillus. Nhiều loài thuộc các chi này có tầm quan trọng thực tế vì chúng tạo ra chất kháng sinh và peptit có hoạt tính chống vi khuẩn, chống vi rút và chống khối u. Chúng cũng tổng hợp các enzym và phân tử có thể điều chỉnh nhiệt có thể ngăn chặn các sinh vật gây bệnh thực vật sinh sống trong đất.
Trong sách chuyên khảo Chi Bacillus , Gordon et al. coi Bt là một loại B. cereus (Bc) cùng với B. anthracis (Ba) và B. mycoides (Bm). Chắc chắn, Bt, Ba và Bc có chung nhiều đặc tính kiểu hình và kiểu gen đến mức cả ba loài được xếp vào một nhóm gọi là Bacillus cereus (BC). Ba là tác nhân gây bệnh than, một bệnh cấp tính và thường gây chết người và động vật. Bc là một tác nhân gây bệnh cơ hội ở người và có thể gây ngộ độc thực phẩm, nhiễm trùng mắt và bệnh nha chu, cùng các bệnh khác. Bt sở hữu một loạt các tính năng đặc biệt bao gồm khả năng sống trong môi trường tự do và không phụ thuộc vào các trực khuẩn Gram dương khác, sản xuất protein tinh thể parasporal entomocidal và tồn tại trong một môi trường độc đáo trong midgut và hemocoel của côn trùng. Ngoài Ba, Bc, Bt và Bm, có hai loài khác có quan hệ họ hàng cao là B. pseudomycoides (Bpm) và B. weihenstephanensi (Bw) trong nhóm BC.
Thật kỳ lạ, Bt đã được cho là một mầm bệnh cơ hội ở động vật và con người. Hai chủng Bt, Bt 97-27 (subsp. Konkukian ) và Bt Al Hakam (được phân lập ở Iraq bởi Ủy ban đặc biệt của Liên hợp quốc), ban đầu được chỉ định là mầm bệnh ở người. Bt 97-27 lần đầu tiên được phân lập từ mô hoại tử ở một bệnh nhân nam 28 tuổi của bệnh viện. Việc chỉ định chủng 97-27 là Bt dựa trên các xét nghiệm sinh hóa và sự xuất hiện của các thể bao gồm. Tuy nhiên, phân lập thứ hai từ cùng một bệnh nhân thiếu các cơ quan thu nhận. Phân tích trình tự cho thấy không có gen diệt côn trùng nào hiện diện trên nhiễm sắc thể 97-27 hoặc một plasmid đơn pBT9727 duy nhất. Phân tích trình tự của bộ gen Al Hakam cũng cho thấy không có ORF được mã hóa nhiễm sắc thể hoặc mã hóa plasmid nào có sự tương đồng đáng kể với bất kỳ gen diệt côn trùng nào. Điều thú vị là pBT9727 của chủng 97-27 và plasmid pXO2 của Ba có các protein sao chép gần như giống hệt nhau và nguồn gốc sao chép được bảo tồn cao. Tuy nhiên, vùng pXO2 mã hóa viên nang axit poly-cD-glutamic đã được thay thế trên pBT9727 bằng các yếu tố di động di truyền, cho thấy rằng pBT9727 có thể đã tiến hóa từ pXO2 để thực hiện các chức năng khác ở chủng 97-27. Vị trí dòng dõi phát sinh loài sử dụng phân tích đa hình chiều dài đoạn khuếch đại (AFLP) và phân tích trình tự so sánh chỉ ra rằng chủng 97-27 liên quan đến Bc và Ba hơn là Bt. Theo hiểu biết của các tác giả, không có nghiên cứu nào khác được báo cáo mô tả Bt là tác nhân gây bệnh cơ hội cho người hoặc động vật máu nóng.
Trình tự bộ gen của một số chủng trong nhóm BC đã được hoàn thiện: Bc (ATCC 14579 và 10987 và chủng E33L), Bw-KBAB4, Ba-Ames, Ba-Ames và Ba-Sterne. Giải trình tự và chú thích bộ gen của Bt subsp. kurstaki sắp hoàn thành trong phòng thí nghiệm Bulla. So sánh tất cả các trình tự bộ gen, bao gồm cả của phân loài kurstaki , cho thấy sự tương đồng rất lớn về nhận dạng trình tự nucleotit và tổ chức gen và operon, một sự kết hợp chưa được quan sát thấy ở đây giữa các loài vi khuẩn khác nhau. Các đặc điểm phân biệt chính của Bt là các yếu tố độc lực và khả năng gây bệnh của nó, được biểu thị bằng các gen diệt côn trùng nằm trên nhiễm sắc thể và một số plasmid. Những chất của Ba là độc tố ba bên của nó và viên nang chỉ được mã hóa bởi plasmid.
Chúng tôi đã xây dựng một cây phát sinh loài, dựa trên sự liên kết trình tự của rRNA 16S của 20 chủng trong nhóm BC bằng cách sử dụng phương pháp nối hàng xóm Bootstrapped (Hình 2). Khối lượng Bootstrap được báo cáo trên các nhánh. Cây hiển thị sự phân chia của 20 chủng thành hai nhóm chính. Nhóm A bao gồm tất cả các chủng Ba, Bc và Bpm và Nhóm B bao gồm chủng Bw KBAB4 và chủng Bm SDANFMO448. Có hai cụm trong Nhóm A, mỗi cụm được đánh dấu bằng số La Mã. Cụm I gồm Ba chủng Sterne CDC684 và Ames A0248; Bc chủng AH187 và AH 820 và ATCC 10987 và 14579; Bpm chủng CIP 5259; và Bt serovar konkukian và Bt Al-Hakm. Bc AH820 và serovar konkukian trong Cụm 1, cả hai đều bị cáo buộc là mầm bệnh ở người, có liên quan chặt chẽ hơn trong khi chủng Bpm CIP 5259 và Bc ATCC 14579, cả hai đều không gây bệnh, là khác biệt nhất trong cụm. Cluster II bao gồm các serovars Bttenebrionis, morrisoni, kurstaki, sotto, israelensis và berliner và Bc G9842. Chúng tôi suy ra từ dendogram rằng cả Bt serovar konkukian và nhóm chủng Al-Hakm khá tốt với các chủng Ba và Bc, nhưng không tốt với các serovars Bt.

Sự biểu hiện và quy định của gen diệt côn trùng
Một trong những khía cạnh ấn tượng nhất của sự hình thành bào tử Bt là sự hình thành các tinh thể cận thị. Độc tố diệt côn trùng (Cry độc tố) của Bt, thường được gọi là δ-endotoxins sau Heimpel, hơi đặc trưng cho một số loài côn trùng nhất định. Họ gen mã hóa các chất độc này là họ gen cry. Đặc điểm chung của các gen cry là chúng được biểu hiện trong giai đoạn tăng trưởng tĩnh. Protein Cry, sản phẩm cuối cùng của sự biểu hiện gen cry, chiếm 20–30% trọng lượng khô của tế bào và thường tích tụ trong tế bào mẹ, bắt đầu từ giai đoạn III của quá trình bào tử và tiếp tục qua giai đoạn VII.
Thuật ngữ δ-endotoxin, liên quan đến độc tố Bt Cry, là một từ nhầm lẫn. Heimpel đã đặt tên cho protein tinh thể parasporal của Bt như vậy vì nó hình thành bên trong tế bào và vì nó đứng thứ tư trong thứ tự các thành phần độc hại khác được phát hiện trong vi khuẩn. Tuy nhiên, nội độc tố có liên quan đến gốc lipo-polysaccharide của phức hợp kháng nguyên soma “O” hoàn chỉnh được tìm thấy trong màng ngoài của các vi khuẩn Gram âm khác nhau và là một yếu tố quan trọng trong khả năng gây bệnh của chúng. Dựa trên phương thức hoạt động của nó, độc tố Cry là một độc tố “đơn giản”, được định nghĩa là một đơn phân hoặc đồng phân của một protein đơn giản độc hại. Độc tố Cry tồn tại dưới dạng một monomer độc hại có khả năng oligome hóa. Các tinh thể parasporal của Bt là các oligome bao gồm các tiểu đơn vị polypeptide protoxin. Protoxin là tiền chất gây độc tức thì của độc tố Cry, tức là khi kích hoạt protoxin, độc tố Cry diệt côn trùng được tạo ra. Điều thú vị là, quá trình xử lý protoxin thành độc tố là khác nhau giữa các nhóm độc tố tương ứng, tùy thuộc vào tính đặc trưng của vật chủ, tức là độc tố giết chết bướm đêm, bọ cánh cứng hoặc muỗi. Ví dụ, độc tố Cry1A và Cry4 (∼65 kDA) chủ yếu tiêu diệt sâu bướm và muỗi, tương ứng, là sản phẩm của protoxin trong phạm vi trọng lượng phân tử 125–135 kDa trong khi độc tố Cry3 (∼68 kDa) giết bọ cánh cứng là sản phẩm chuyển đổi trong số các protoxin 72-kDa. Cơ chế hoạt động của Cry toxin sẽ được thảo luận sau trong bài tổng quan này.
Mức độ tổng hợp protein Cry cao dường như được kiểm soát đồng thời bởi nhiều cơ chế khác nhau xảy ra ở cấp độ phiên mã và sau phiên mã. Thông thường, sự khởi đầu của quá trình tạo bào tử ở Bt được kiểm soát bằng cách kích hoạt liên tiếp các yếu tố sigma liên kết với RNA polymerase lõi và phiên mã trực tiếp sử dụng các promoter đặc hiệu tạo bào tử. Có năm yếu tố phiên mã đặc hiệu cho quá trình tạo bào tử, σ H , σ F , σ E , σ G và σ K , được điều chỉnh chặt chẽ và xuất hiện theo thứ tự cố định trong quá trình tạo bào tử. Yếu tố σ H đang hoạt động trong ô tiền phân chia; σ E và σ K đang hoạt động trong tế bào mẹ; và σ F và σ G đang hoạt động trong tiền nghiệm. Một số chất xúc tiến gen khóc đã được xác định và xác định trình tự của chúng. Các cry1Aa gen, ví dụ, được thể hiện qua hai quảng bá chồng chéo (BTI và BtII). Transcription từ promoter hạ lưu, BTI, đang hoạt động giữa các giai đoạn II và VI của hình thành bào tử và sử dụng σ E . Quá trình phiên mã sau đó được tạo điều kiện thuận lợi bởi chất xúc tiến ngược dòng, BtII, hoạt động từ giai đoạn VI đến cuối quá trình bào tử và sử dụng σ K. Việc sử dụng tuần tự hai trình tự khởi động rất có thể đảm bảo tổng hợp protoxin trong suốt chu kỳ bào tử. Tuy nhiên, một số gen khóc dường như được biểu hiện độc lập với sự hình thành bào tử. Ví dụ, biểu hiện gen cry3A được tăng cường và kéo dài ở các chủng đột biến không thể bắt đầu hình thành bào tử. 66 Ngoài ra, De Souze và cộng sự. và Malvar et al. 68 báo cáo rằng gen cry3A được biểu hiện trong quá trình sinh dưỡng. Các cry3A promoter, được công nhận bởi σ Một , yếu tố sigma của các tế bào sinh dưỡng, được kích hoạt ở phần cuối của sự tăng trưởng sinh dưỡng mũ và vẫn hoạt động thông qua giai đoạn III của hình thành bào tử.
Một cơ chế điều hòa bổ sung để biểu hiện một số gen kêu ở cấp độ phiên mã liên quan đến một protein điều hòa liên kết với các ái lực khác nhau trong một lần lặp lại đảo ngược và một vùng uốn cong tiềm ẩn nằm 200–300 cặp bazơ ngược dòng với các chất xúc tiến cry1 khác nhau. Protein điều hòa này được xác định là tiểu đơn vị E2 của pyruvate dehydrogenase. Một chimera bao gồm vùng ngược dòng bị đột biến và trình tự mã hóa của β-galactosidase dẫn đến giảm liên kết của tiểu đơn vị E2 và giảm tổng hợp β-galactosidase. Rõ ràng, sự tham gia của tiểu đơn vị E2 của pyruvate dehydrogenase trong quy định gen khóc có liên quan đến mối liên hệ giữa hoạt động dị hóa và tổng hợp độc tố Cry.
Đọc tiếp tại: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3035146/
Các từ viết tắt
| AC | adenylyl cyclase |
| AFLP | tính đa hình chiều dài đoạn khuếch đại |
| BC | Nhóm Bacillus cereus |
| Bt | Bacillus thuringiensis |
| ORF | khung đọc mở |
| PKA | protein kinase A |
| TBR | vùng liên kết độc tố |